Des populations structurées
Les taux démographiques des individus déterminent le taux de croissance des populations (Caswell, 2001). Or, les individus au sein d’une population peuvent différer dans leurs performances en fonction de traits tels que l’âge et le sexe (Caswell, 2001). Ces hétérogénéités entre groupes d’individus définissent la structure d’une population. Une autre caractéristique importante d’une population est sa densité ou sa taille. En effet, la densité dépendance est l’un des principaux mécanismes de régulation des populations animales et végétales (Sibly et al., 2005, Henle et al., 2004).
La structure des populations évolue dans le temps et dans l’espace sous l’influence des réponses individuelles au sein des classes d’âge et de sexe aux variations de l’environnement abiotique (ex : climat, Coulson et al., 2001), trophique (ex : densité de proies et de prédateurs, de Roos et al., 2003), et social (ex : nombre de compétiteurs et de partenaires sexuels, Begon et al., 2006, Gascoigne et al., 2009). Ainsi, il existe un lien de rétroaction fort entre la structure des populations et l’histoire de vie, la structure des populations influençant les performances des individus à travers les interactions sociales et les performances des individus déterminant la structure de population. Ici, nous nous concentrerons sur le lien existant entre la structure de population, l’histoire de vie et la dynamique de petites populations isolées. Nous ne discuterons donc pas de la structuration spatiale des populations.
Structure de population et histoire de vie
Structure d’âge et effets cohortes
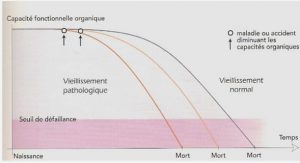
Dans les populations structurées en cohortes de naissance, c’est-à-dire en groupes d’individus nés pendant un même épisode de reproduction, des effets cohortes émergent lorsque la variation environnementale génère de la variation d’histoire de vie entre cohortes (Albon et al., 1987, Gaillard et al., 2003, Lindström, 1999). Deux types d’effets cohortes ont été décrits par les écologistes des populations : les effets cohortes numériques directs et les effets cohortes qualitatifs décalés. La variation environnementale produit des effets cohortes numériques directs lorsqu’elle influence la survie et les trajectoires de croissance des individus avant la maturité sexuelle et donc provoque de la variation dans le taux de recrutement (ex : chez les ongulés, Gaillard et al., 2003). Les effets cohortes qualitatifs décalés sont quant à eux la conséquence des effets à long terme des conditions environnementales sur les performances et la qualité intrinsèque des individus et peuvent être transmis entre générations via des effets parentaux (ex : Marquis et al., 2008, Plaistow et al., 2006). Les effets à long terme de l’environnement peuvent ainsi générer des effets « cuillère en argent » (ou « silver spoon effects », Grafen, 1988), lorsque des conditions favorables confèrent un avantage durable de valeur sélective aux individus par rapport à des individus ayant expérimenté des conditions défavorables. De tels effets de l’environnement créent de l’autocorrélation individuelle positive des traits d’histoire de vie (ex : taux de croissance corporelle plus élevé tout au long de la vie chez des espèces à croissance indéterminée, Madsen & Shine, 2000) et peuvent générer des covariations positives entre traits d’histoire de vie (ex : entre la survie et la reproduction, van de Pol et al., 2006). Cependant, des phénomènes de compensation de type croissance compensatoire, lorsque les conditions environnementales s’améliorent, peuvent également produire des covariations négatives entre les traits, ces phénomènes étant souvent associés à des coûts physiologiques et/ou écologiques (ex : augmentation du taux métabolique et du risque de prédation, Metcalfe & Monaghan, 2001).
Les conditions climatiques, la densité de population, la disponibilité en nourriture et la qualité de l’habitat sont les principaux facteurs environnementaux à l’origine d’effets cohortes (ex : Albon et al., 1987, Forchhammer et al., 2001, Reid et al., 2003, Roach, 2003). Les conditions environnementales pendant le développement précoce sont souvent prédites comme étant à l’origine des effets cohortes (Lindström, 1999, Metcalfe & Monaghan, 2001). Cependant, les conditions expérimentées plus tard dans la vie peuvent également générer de la variation cohorte, notamment par l’intermédiaire d’effets intergénérationnels des conditions expérimentées par la mère pendant la saison de reproduction (ex : Marquis et al., 2008, Plaistow et al., 2006). Les différentes routes d’influence de l’environnement sur l’histoire de vie pouvant générer des effets cohortes numériques et qualitatifs sont discutées en détail Chapitre 1 et illustrées à l’aide d’exemples trouvés dans la littérature. Elles comprennent (i) les effets directs de l’environnement sur la qualité des individus au sein des cohortes tels que présentés précédemment, c’est-à-dire des effets densité indépendants, (ii) ainsi que ses effets indirects via l’intensité de la compétition intraspécifique intra- et inter-cohortes, c’est-à-dire des effets densité-dépendants. Deux types de compétitions intraspécifiques peuvent être distingués. Les effets cohortes densité-dépendants peuvent impliquer des interactions sociales de types symétriques, c’est-à-dire entre individus d’aptitudes compétitrices équivalentes, au sein d’une même cohorte (compétition intra-cohortes) ou entre cohortes (compétition intercohortes Beckerman et al., 2003, Lindström & Kokko, 2002). Cependant, la compétitivité des individus étant fortement influencée par leur taille (ou masse) corporelle (Bystrom & Andersson, 2005, Kooijman, 2000, Persson et al., 1998), les effets cohortes densitédépendants impliquent généralement des interactions sociales asymétriques entre individus appartenant à différentes cohortes, de types compétition par exploitation asymétrique des ressources et cannibalisme (ex : Claessen et al., 2000, Claessen et al., 2002, Persson et al., 1998). Dans ce dernier cas nous verrons que la structure d’âge de la population peut avoir des effets importants sur les trajectoires d’histoire de vie des cohortes en compétition (voir Chapitre 1 et Claessen et al., 2000, Claessen et al., 2002, Persson et al., 1998).
Densité de population
La densité (ou la taille) de population peut affecter les performances des individus de manière négative (ex : Forrester et al., 2011, Massot et al., 1992, Bonenfant et al., 2009) ou positive (ex : Angulo et al., 2007, Berec et al., 2001, Courchamp et al., 2000). La densité dépendance négative est le résultat de la compétition intraspécifique pour l’accès à des ressources limitées telles que la nourriture ou l’espace (Begon et al., 2006). Elle peut être due à des interactions indirectes entre compétiteurs via l’exploitation et la déplétion des ressources, c’est-à-dire de la compétition par exploitation, ou à des interactions directes entre individus via des phénomènes d’interférence, c’est-à-dire de la compétition par interférence, où les meilleurs compétiteurs monopolisent les ressources (Begon et al., 2006). A l’opposé, la densité dépendance positive, aussi appelée effet Allee (Allee et al., 1949), est due à la réduction du nombre d’interactions sociales coopératives à faibles densités telles que la rencontre de partenaires sexuels potentiels, la facilitation de la reproduction, la reproduction coopérative ou encore la dilution de la prédation (voir Chapitre 2 dans Courchamp et al., 2008). Deux types d’effets Allee peuvent être distingués : les effets Allee élémentaires ou « component Allee effects » et les effets Allee démographiques ou « demographic Allee effects » (Stephens et al., 1999). Les effets Allee élémentaires correspondent à de la densité dépendance positive sur une des composantes de la valeur sélective des individus, par exemple sur la survie, tandis que les effets Allee démographiques correspondent à de la densité dépendance positive sur la valeur sélective globale des individus, qui dépend à la fois de paramètres de survie et de paramètres de reproduction (Courchamp et al., 2008). Ainsi, un effet Allee élémentaire ne se traduit pas nécessairement par une réduction de la valeur sélective des individus à faibles densités et donc par un effet Allee démographique s’il existe par exemple des compromis entre les différentes composantes de la valeur sélective tels qu’une compensation des coûts de survie à faible densité par une fécondité plus élevée (Stephens et al., 1999, Courchamp et al., 2008, Gascoigne et al., 2009).
La détection et la caractérisation de la forme de la densité dépendance de l’histoire de vie est souvent rendue difficile par le manque de données empiriques de bonne qualité et couvrant une large gamme de densités de population, ce qui est particulièrement vrai pour les populations en déclin (Morris & Doak, 2002, Courchamp et al., 2008, Beissinger & McCullough, 2002, Ferrière et al., 2004). Cependant, une relation négative entre la densité de population et la survie, la reproduction, la croissance, et des traits physiologiques tels que l’immunocompétence a été décrite chez un grand nombre de taxa (ex : chez les poissons, Edeline et al., 2010, Forrester et al., 2011, les mammifères, Bonenfant et al., 2009, Koskela et al., 1999 , les oiseaux, Both, 1998, les arthropodes, , les reptiles, Massot et al., 1992, Svensson et al., 2001 , et les plantes Turkington et al., 2005). Des effets Allee élémentaires ont également été décrits chez un grand nombre d’espèces (voir revue par Courchamp et al. (2008) chez des mammifères, oiseaux, insectes, reptiles, amphibiens, poissons et autres organismes marins). En revanche, il n’existe que peu d’exemples empiriques d’effets Allee démographiques (voir revue par Gregory et al. , 2010). Ainsi, alors que des effets Allee élémentaires provoqués par un manque de partenaires sexuels ont été rapportés chez une grande variété d’espèces marines et terrestres, des évidences empiriques d’effets Allee démographiques n’ont été décrites que chez deux espèces de papillons, une espèce de crustacée et une espèce de plante (Gascoigne et al., 2009). Une approche alternative pour lier la densité à un effet Allee démographique, lorsqu’un effet Allee élémentaire a été détecté, est de combiner des données empiriques d’histoire de vie avec des modèles populationnels à deux sexes incluant des fluctuations du sexe ratio et une fonction d’appariement décrivant la formation des couples reproducteurs (ex : Bessa-Gomez et al., 2004, Engen et al., 2003, Legendre et al., 1999).
Structure de sexe
La structure de sexe d’une population peut affecter de manière importante la valeur sélective des mâles et des femelles. En effet, elle détermine le taux de fertilisation des femelles reproductrices (Rankin & Kokko, 2007) et influence fortement l’accès à la reproduction des mâles (ex : Mysterud et al., 2003, Fitze et al., 2008). La théorie sur l’évolution du sexe ratio prédit que les individus du sexe le plus rare ont une valeur sélective plus élevée que les individus du sexe le plus abondant (ex : Fisher, 1930). Le sexe ratio adulte devrait donc être fortement régulé par la compétition intersexuelle et varier faiblement dans le temps et dans l’espace. Cependant, la variation des conditions environnementales peut générer des fluctuations dans la structure de sexe si les mâles et les femelles ne répondent pas de la même manière à l’environnement (ex : Coulson et al., 2001, Tschirren et al., 2003, Metcalfe & Monaghan, 2001 , Le Galliard et al., 2005d). C’est particulièrement le cas chez les espèces présentant un dimorphisme sexuel de taille corporelle, où le sexe le plus grand fait face à des demandes énergétiques plus élevées (Lindström, 1999, Coulson et al., 2001 , Clutton-Brock et al., 1985). Ainsi, la théorie sur l’allocation optimale au sexe (« optimal sex allocation theory », Charnov, 1982) prédit que les variations du sexe ratio générées par l’environnement devraient être compensées par une augmentation de l’investissement maternel dans le sexe rare. Cependant, chez des espèces où la détermination du sexe est génétique (chez les plupart des vertébrés) et les soins parentaux sont limités ou inexistants, les capacités des femelles à ajuster leur investissement en fonction du sexe de leurs descendants peuvent être réduites (Cockburn et al., 2002). Par exemple, chez une espèce polygyne où les comportements sexuels des mâles sont délétères pour la survie et la fécondité des femelles, une incapacité des femelles à ajuster le sexe ratio primaire en réponse à un sexe ratio adulte mâle-biaisé peut conduire à une augmentation du biais de sexe ratio vers les mâles, réduisant d’autant plus la valeur sélective des femelles (ex : Le Galliard et al., 2005c, Le Galliard et al., 2005d).
|
Table des matières
Introduction générale
Contexte de l’étude
Influences de la structure de population sur l’histoire de vie du lézard vivipare
Influences de la structure de population sur le système d’appariement et l’intensité de la sélection sexuelle chez le lézard vivipare
Conséquences pour la dynamique des populations de lézards vivipares
Conclusion
Références
Annexes
Chapitre 1 – Effets cohortes dans des populations structurées et conséquences démographiques
Chapitre 2 – Effets de l’environnement nutritif précoce sur l’histoire de vie dans des populations structurées en cohortes
Chapitre 3 – Effets de l’environnement nutritif tardif sur l’histoire de vie dans des populations structurées en cohortes
Chapitre 4 – Densité de population et interactions trophiques proies-prédateurs
Chapitre 5 – Densité dépendance de l’histoire de vie et risque d’extinction
![]() Télécharger le rapport complet
Télécharger le rapport complet